Атф синтаза у кишечной палочки
Строение и функционирование F1F0-АТФ-синтазы
Основная масса энергии в клетках эукариот и почти всех прокариот запасается в процессе окислительного фосфорилирования в форме АТФ. Этот многоступенчатый процесс происходит в мембранах.
В организме человека весом около 70 кг за 75 лет при сидячем образе жизни нарабатывается 2 млн кг АТФ. В клетках эукариот синтез АТФ, который является одним из наиболее распространенных ферментативных биологических процессов, происходит в специальных органеллах, называемых митохондриями.
Образование АТФ в клетке катализируется ферментом F1F0-АГФ-синтазой, который представляет собой необычный молекулярный мотор, использующий энергию электрохимического градиента протонов (трансмембранный протонный градиент) для синтеза АТФ. Электрохимический градиент протонов создается комплексами, переносящими электроны в ходе окислительного фосфорилирования.
Он определяется двумя показателями: мембранным потенциалом и разностью концентраций протонов по сторонам мембраны митохондрий.
Общая структура F1F0-АТФ-синтазы во всех клетках одинакова, хотя состав субъединиц может различаться. Простейшая форма F1F0-АТФ синтазы цитоплазматической мембраны бактериальной клетки состоит из восьми типов субъединиц и обладает молекулярной массой 530 кДа.
Состав ее может быть выражен стехиометрической формулой а3 b3 γ δ ε a b2 с10-14. В митохондриях фермент содержит 7-9 дополнительных регуляторных субъединиц, которые немного увеличивают его молекулярную массу. АТФ-синтаза состоит из двух доменов:
• связанный с мембраной домен F0 (ab2c10-14 у бактерий), который участвует в транслокации протонов в направлении электрохимического градиента, и
• глобулярный домен F1(а3 β3 γ δ ε у бактерий), содержащий каталитические сайты, ответственные за синтез АТФ. (Изолированный F1 домен обладает способностью к гидролизу АТФ.)
Субъединицы с домена F0 образуют кольцо, которое взаимодействует с субъединицей а. Субъединица у формирует центральный выступ ротора, который связан с основанием кольца с и входит в каталитический домен F1. Этот домен состоит из трех а и трех b субъединиц, которые расположены попеременно и образуют структуру гексагонального цилиндра вокруг субъединицы у.
Выступающая ножка статора состоит из субъединиц b2δ, причем субъединица δ связана с доменом F1, а b2 с доменом F0 митохондриальной мембраны и с субъединицей а.
Каким образом F1F0-АТФ-синтаза использует энергию трансмембранного протонового градиента для синтеза АТФ? Предложенная модель постулирует существование следующих основных этапов:
• субъединицы а и с контролируют транспорт протонов таким образом, что с-кольцо вращается по отношению к субъединице а. Иными словами, энергия электрохимического градиента протонов расходуется на механическое вращение субъединиц с;
• у-субъединица центрального выступа вращается вместе с с-кольцом, при этом трансмембранная движущая сила протонов передается на расстояние 100 А домен F1; и
• механическая энергия вращения используется для высвобождения АТФ, синтез которого катализируется b-субъединицами домена F1.
Поэтому вращение с-кольца и центральной у-субъединицы относительно субдомена а3 b3 играет существенную роль в сопряжении трансмембранной движущей силы протонов с образованием и выходом АТФ. Поскольку каждая с-субъединица несет один протон, то при полном обороте с-кольца (в зависимости от типа АТФазы) транспортируются 10-14 протонов, и на одну молекулу образующегося АТФ транспортируются около четырех протонов.
F1F0-АТФ-синтаза превращает электрохимическую энергию в механическую и снова в химическую, почти со 100%-ной эффективностью. Синтез АТФ может проходить с максимальной скоростью порядка 100 с-1, и уровень АТФ в клетке может достигать миллимолярных концентраций.
У некоторых бактерий F1F0-АТФ-синтаза работает в обратном направлении, и энергия, высвобождающаяся за счет гидролиза АТФ, расходуется на выход протонов из клетки и на создание их градиента по сторонам цитоплазматической мембраны. Затем энергия протонного градиента используется для транспорта в клетку метаболитов, например лактозы.
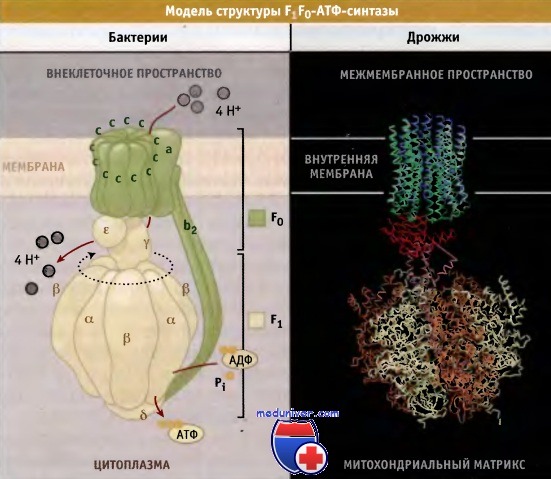
Схематическое изображение комплекса F1F0-АТФ-синтазы Е. coli и часть кристаллической структуры дрожжевого фермента.
Изображение кристаллической структуры дрожжевого фермента построено на основании Protein Data Bank file 1Q01.
Показано предполагаемое положение мембраны.
– Также рекомендуем “Строение и функционирование протонных насосов (H+-АТФазы, V-АТФазы)”
Оглавление темы “Биология клетки”:
- Строение и функционирование натрий-калиевой АТФазы
- Строение и функционирование F1F0-АТФ-синтазы
- Строение и функционирование протонных насосов (H+-АТФазы, V-АТФазы)
- Современные возможности изучения ионных каналов
- Вывод и применение уравнения Нернста
- Механизмы входящего выпрямления калиевых каналов мембраны клетки
- Развитие муковисцидоза (кистозного фиброза) при мутации гена анионного канала CFTR
- Секрекция и адресование белков в клетке
- Адресование (таргетинг) белков в эндоплазматическом ретикулуме клетки
- Сигнальные последовательности связывания белков с эндоплазматическим ретикулумом (ЭПР)
Источник
АТФ-синтаза — фермент (КФ 3.6.3.14), который осуществляет реакцию синтеза АТФ из АДФ и аниона фосфата обычно за счет энергии трансмембранного электрохимического потенциала протонов (то есть комбинации градиента протонов и электрического напряжения), а в некоторых организмов за счет электрохимического потенциала ионов натрия , превращая ее, таким образом, в энергию химических связей, которая затем может использоваться клеткой в биохимических реакциях. В случае, когда фермент проводит обратный процесс — формирует трансмембранный протонный градиент за счет гидролиза АТФ, его могут называть АТФазы. Действие фермента ингибирует антибиотик олигомицин.
Номенклатура
АТФ-синтаза F 1 F 0 состоит из двух субъединиц:
- F 0 мембранная часть комплекса
- F 1 часть комплекса в матриксе митохондрий или цитоплазме бактерий.
Номенклатура субъединиц АТФ-синтазы достаточно сложная и имеет долгую историю. F 1 -фракции получила свое название от срока fraction 1 («фракция 1»), а F 0 (первоначально писалась с индексом «O», а не «ноль», как это более принято сейчас) получила название как фракция, н “связывает олигомицин.
По примеру других ферментов, большинство субъединиц получили названия в виде греческих (от α до ε) и латинских (от a до h) букв. Остальные субъединиц получили сложные названия:
- F 6 (отfraction 6)
- OSCP (oligomycin sensitivity conferral protein — «вспомогательной белок чувствительности к олигомицину»)
- A6L (названный по названию гена генома митохондрий, кодирующего эту субъединицу)
- IF1 (inhibitory factor 1 — «ингибирующее фактор 1»)
Устройство и принцип работы
АТФ-синтаза располагается на одной из мембран клетки и состоит из погруженного в нее домена F 0 и выступающего в матрикс или цитоплазму домена F 1, соединенных субъединицей γ. Отдаленно фермент напоминает плодовое тело гриба (в связи с чем в литературе по клеточной биологии, особенно старой, АТФ-синтазу иногда называли «грибовидными телом»).
Комплекс F 1 имеет диаметр около 9-10 нм и при разрушении γ «ножки» может отделяться от мембраны, образуя т F 1 -АТФазу. F 1 состоит из трех субъединиц α и трех β, которые объединяются попарно и формируют гексамеров с тремя активными центрами. Конформации меняются при вращении субъединицы γ вместе с комплексом F 0. Движущей силой в этом процессе переноса протона, что катализирует доменом F 0. Таким образом, протон напрямую не участвует в реакции конденсации АДФ и аниона фосфата. Следует отметить, что сам α 3 β 3 гексамеров не вращается относительно «статора» a, поскольку он содержится субъединицей δ, в свою очередь связанной с a субъединицей b («стеблем») комплекса F 1 (хотя обычно F 1 рассматривается как неподвижный , в действительности оба комплекса вращаются относительно друг друга в противоположных направлениях).
Принцип работы комплекса F 1 состоит сначала в слабом связывании АДФ и фосфата с активным центром, который затем меняет конформацию и прочно связывает их, в результате чего синтез АТФ идет самопроизвольно. При третьей конформации АТФ выталкивается из активного центра.
Принцип работы комплекса F 0 (что часто называют «самым маленьким в мире роторным электромотором») заключается в проникновении протона через канал в «статоре» (субъединица a) до связывания в «Ротор» (c-кольце). Для освобождения на другой стороне мембраны протона нужно выйти через другой канал в статоре, что сдвинут на некоторое расстояние, то есть для выхода ротора НЕОБХОДИМО обернуться относительно статора. Таким образом в роторе создается горизонтальная разность потенциалов, вращает его относительно статора.
Физиологическая роль
В роли АТФазы фермент применяется анаеробними бактериями для создания за счет энергии АТФ трансмембранного электрохимического потенциала протонов. Этот градиент, в свою очередь, используется для вращения жгутиков и для транспорта ионов внутрь клетки.
В аэробных бактерий фермент в основном используется для синтеза АТФ, причем электрохимического потенциал для этого производится при функционировании дыхательная цепь переноса электронов. В целом данный процесс называется окислительным фосфорилированием. Он протекает в митохондриях эукариот, на внутренней мембране которых расположены молекулы АТФ-синтазы, причем F 1 субъединица находится в матриксе, где и протекает процесс синтеза АТФ из АДФ и фосфата.
АТФ-синтеза задействована также в процессе фотосинтеза; она локализуется на тилакоидних мембранах хлоропластов, ориентируясь F 1 субъединицы в строму. Строение и механизм работы фермента в этом случае практически идентичен таковым для АТФ-синтазы митохондрий, однако протонный электрохимический потенциал формируется в принципиально ином электронтранспортной цепи.
Изображения по теме
Источник
АДФ + PO43- + Е ⇔ АТФ
АТФ используется клетками как источник энергии во многих клеточных процессах. Та же самая реакция может идти и в обратном направлении, когда АТФ расщепляется на специальном белке-катализаторе обратно на АДФ и фосфат с выделением энергии.
АТФ-синтаза состоит из двух механизмов. Первый, F0, это электромотор, находящийся в клеточной мембране и превращающий энергию, запасенную в разности потенциалов по разные стороны клеточной мембраны. Липидная мембрана служит изолятором в этой электрохимической «батарейке»: через нее ионы не проходят. Разность потенциалов создается другими сложными механизмами в конечном счете из «сжигания» сахара в кислороде. Ион водорода H+ втягивается во «впускной коллектор» и присоединяется к белковой дольке ротора. Ротор поворачивается за счет электростатических сил, а долька, достигшая «выхлопного коллектора» мотора, освобождается от иона каталитическим белком, и этот ион проваливается внутрь клетки, опять же за счет электростатических сил, стремящихся выровнять потенциал по обе стороны мембраны. Таким образом, электроэнергия сначала превращается в механическую энергию вращения молекулярного вала, присоединенного к ротору и уходящего вглубь клетки, к механизму синтеза, F1.
Механико-химический реактор F1 состоит из трех белковых долек, каждая из которых состоит из двух белковых молекул (их называют α-F1 и β-F1, а вал сделан из одной молекулы, обозначаемой γ-F1). Каждая долька может принимать две устойчивые пространственные конфигурации за счет взаимного межатомного притяжения — как обычный настенный выключатель оказывается в двух устойчивых положениях, хотя промежуточные положения неустойчивы. Одно из этих положений, однако, имеет более высокую энергию. Молекулы сдвигаются в конфигурацию с более высокой энергией за счет асимметрии вращающегося γ-вала, как будто бы «кулачком» на нем.
Когда к αβ-комплексу присоединяется АДФ и ион фосфата, равновесие нарушается, и молекула, как пружинка с запасенной энергией, перепрыгивает в состояние с меньшей энергией, а запасенная энергия тратится на сближение АДФ и фосфатного иона, в результате чего те соединяются в молекулу АТФ, в конечном счете уносящую этот запас энергии.
Вращение механизма можно увидеть в микроскоп, если присоединить к ротору в F0 специально изготовленную длинную светящуюся (флюоресцирующую) молекулу-стержень. В самом конце фильма можно увидеть реконструкцию этого потрясающего опыта Масасуке Ёсиды и врезку с данными, показывающими вращение ротора.
Интересно, что на нижнем конце ротора имеется еще один белок, δ-F1, который тоже умеет изменять конфигурацию в присутствии АДФ, исходного реагента для реакции. Когда АДФ вокруг реактора оказывается мало, этот белок меняет форму и заклинивает ротор, чтобы не расходовать электрохимическую энергию вхолостую, поскольку продвижение ионов H+ через остановленный ротор невозможно. Да, и об экономии клеточной электроэнергии природа тоже «подумала»!
Больше информации о работе АТФ-синазы можно найти в конспекте А. Крафтса из университета Иллинойса (англ.), а кое-что из истории исследования этого механизма здесь, по-русски.
Источник
Биологическая химияЛелевич Владимир Валерьянович
Строение АТФ-синтазы
АТФ-синтаза – интегральный белок внутренней мембраны митохондрий. Он расположен в непосредственной близости к дыхательной цепи и обозначается как V комплекс. АТФ-синтаза состоит из 2 субъединиц, обозначаемых как F0 и F1. Гидрофобный комплекс F0 погружен во внутреннюю мембрану митохондрий и состоит из нескольких протомеров, образующих канал по которому протоны переносятся в матрикс. Субъединица F1 выступает в митохондриальный матрикс и состоит из 9 протомеров. Причем три из них связывают субъединицы F0 и F1, образуя своеобразную ножку и являются чувствительными к олигомицину.
Суть хемиоосмотической теории: за счет энергии переноса электронов по ЦТД происходит движение протонов через внутреннюю митохондриальную мембрану в межмембранное пространство, где создается электрохимический потенциал (??Н+), который приводит к конформационной престройке активного центра АТФ-синтазы, в результате чего становится возможным обратный транспорт протонов через протонные каналы АТФ-синтазы. При возвращении протонов назад электрохимический потенциал трансформируется в энергию макроэргической связи АТФ. Образовавшаяся АТФ с помощью белка-переносчика транслоказы перемещается в цитозоль клетки, а взамен в матрикс поступают АДФ и Фн.
Коэффициент фосфорилирования (Р/О) – число атомов неорганического фосфата, включенных в молекулы АТФ, в пересчете на один атом использованного поглощенного кислорода.
Пункты фосфорилирования – участки в дыхательной цепи, где энергия транспорта электоронов используется на генерацию протонного градиента, а затем в ходе фосфорилирования запасается в форме АТФ:
1. 1 пункт – между пиридинзависимой и флавинзависимой дегидрогеназами; 2 пункт – между цитохромами b и с1; 3 пункт – между цитохромами а и а3.
2. Следовательно, при окислении НАД-зависимых субстратов коэффициент Р/О равен 3, так как электроны от НАДН транспортируются с участием всех комплексов ЦТД. Окисление ФАД-зависимых субстратов идет в обход I комплекса дыхательной цепи и Р/О равен 2.
Данный текст является ознакомительным фрагментом.
Похожие главы из других книг:
Строение
Колибри образуют весьма цельное семейство Trochilidae. Они малы; самые крупные – не больше ласточки, самые мелкие – крошечные создания, не крупнее шмеля. Они отличаются очень коротенькими ножками и маленькими лапками, очень длинными острыми крыльями, длинным тонким
V. Состав и строение живых тел
Наблюдая жизнь растений, животных и человека, мы видим, что с ними непрестанно совершаются самые разнообразные перемены: они растут, размножаются, стареют, умирают. Внутри них постоянно движутся разные соки, газы, пища и пр. Поступающие в
Строение ВТМ
Вирус табачной мозаики изучен гораздо лучше других вирусов. Его размеры и форма нам уже хорошо известны. Но каково внутреннее строение частицы этого вируса?Рибонуклеиновая кислота находится в вирусной частице в виде простых спиралей, которые скрыты в
КЛЕТОЧНОЕ СТРОЕНИЕ ОРГАНИЗМОВ СТРОЕНИЕ КЛЕТКИ. ПРИБОРЫ ДЛЯ ИССЛЕДОВАНИЯ СТРОЕНИЯ КЛЕТКИ
1. Выберите один наиболее правильный ответ.Клетка – это:A. Мельчайшая частица всего живогоБ. Мельчайшая частица живого растенияB. Часть растенияГ. Искусственно созданная единица для
9.1. Строение пищеварительного канала
Пищеварительный канал состоит из системы органов, которые производят механическую и химическую обработку пищи и ее всасывание. У человека пищеварительный канал имеет вид трубки длиной 8-10 м. Стенка пищеварительной трубки состоит из
2. НЕЙРОН. ЕГО СТРОЕНИЕ И ФУНКЦИИ
Мозг человека состоит из 10 12 нервных клеток. Обычная нервная клетка получает информацию от сотен и тысяч других клеток и передаёт сотням и тысячам, а количество соединений в головном мозге превышает 10 14 – 10 15 . Открытые более 150 лет тому назад
2. Строение и жизнь микробов
Итак, микробами называются мельчайшие живые организмы, невидимые простым глазом. Как мы узнали из предыдущей главы, к микробам относятся разнообразные формы живой материи, имеющие как клеточную, так и неклеточную организацию.Различают
Глава 1. Строение мозга
Мозг человека – самый сложный и загадочный орган тела. Для того чтобы его понять, человечеству пришлось потратить несколько столетий, но чем дальше мы продвигаемся в изучении, тем больше возникает новых вопросов. По большому счету, мозг
Строение зуба
Зубы – органы ротовой полости, выполняющие различные функции, главной из которых является механическая обработка корма. Общий план структуры зубов характерен для обеих генераций (молочной и постоянной). Анатомически в зубе различают коронку, шейку и
История, легенда и строение
Бомбилья, она же бомбижья, бомбиша, момбиша, бомба (в Бразилии), масасса (в Сирии и Ливане) – это трубочка, через которую пьют мате. Бомбилья может быть бамбуковой, тростниковой, одноразовой пластмассовой, но чаще всего она металлическая. В
Источник