Кишечная палочка клеточная стенка
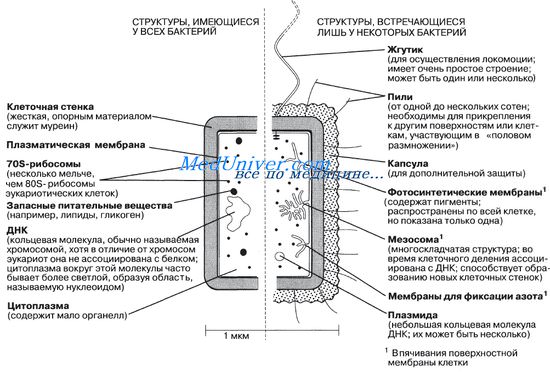
Оглавление темы “Строение бактерий. Размножение бактерий.”: Строение бактерий. Клеточная стенка бактерий.На рисунке показано строение обобщенной бактерии — типичной прокариотической клетки. На рисунке А изображена широко известная палочковидная бактерия Escherichia coli. Обычно она совершенно безвредна. Ее наличие в воде может использоваться в качестве очень надежного показателя загрязнения воды фекалиями. Из всех бактерий E.coli изучена лучше всего. Кроме того, это одна из бактерий, генетическая карта которых установлена полностью. Обратите внимание, что у Е. coli намного меньше видимых внутриклеточных структур, чем в эукариотиче-ской клетке (рис. 5.10 и 5.11). На рис. 2.7 показана другая палочковидная бактерия, у которой в отличие от E.coli имеется жгутик.
Клеточная стенка бактерийКлеточная стенка бактерий — структура довольно прочная и позволяет клетке сохранять свою форму; это обусловлено наличием в ней муреина — молекулы, построенной из параллельных полисахаридных цепей, перекрестно связанных через регулярные интервалы короткими цепями аминокислот. Таким образом, каждая клетка окружена как бы сетчатым мешком, представляющим на деле одну огромную молекулу. Клеточная стенка предохраняет клетку от разрыва при поступлении в нее воды (например, в результате осмоса). Ионы воды и малые молекулы попадают в клетку через мельчайшие поры в клеточной стенке. В 1884 г. датский биолог Кристиан Грам разработал метод окрашивания, с помощью которого было установлено, что бактерии подразделяются на две естественные группы, что, как теперь стало известно, обусловлено различиями в строении их клеточной стенки. Одни бактерии, окрашивающиеся по Граму, получили название грамположительных, другие, не окрашивающиеся, — грамотрицательных.
У грамположительных бактерий, таких как Staphylococcus, Bacillus и Lactobacillus в муреиновую сетку встроены другие компоненты, в основном полисахариды и белки, что делает клеточную стенку сравнительно толстой. У грамотрицательных бактерий, таких как Salmonella, E.coli и Azotobacter, клеточная стенка тоньше и имеет более сложное строение (рис. 2.8). Муреиновый слой у этих бактерий снаружи покрыт гладким тонким мембраноподобным слоем липидов и полисахаридов, защищающим клетки от лизоцима — антибактериального фермента, содержащегося в слезах, слюне и других биологических жидкостях, а также в белке куриного яйца. Лизоцим расщепляет полисахаридный каркас муреина, что приводит к продырявливанию клеточной стенки и лизису клетки, т. е. к ее осмотическому набуханию и разрыву. Липидно-полисахаридный слой обусловливает также устойчивость грамот-рицательных бактерий к пенициллину. Этот антибиотик блокирует образование перекрестных сшивок в муреине растущих грамположительных бактерий, что делает их клетки более чувствительными к осмотическому шоку. Видео строение бактериальной клетки – анатомия бактерии– Также рекомендуем “Плазматическая мембрана. Мезосомы. Хромосома. Рибосомы.” |
Источник
Оглавление темы “Анатомия бактериальной клетки. Физиология бактерий.”:
1. Анатомия бактериальной клетки. Поверхностные структуры бактерии. Капсула бактерий. Организация капсул. Окраска капсул бактерий. Состав капсул. Антигенные свойства капсул.
2. Жгутики бактерий. Расположение жгутиков. Перитрихи. Монотрихи. Политрихи. Лофотрихи. Амфитрихи. Феномен роения. Диагностика подвижности бактерий.
3. Микроворсинки бактерий. Фимбрии бактерий. F-пили ( секс-пили ) бактерии. Клеточная оболочка бактерий. Гликокаликс.
4. Клеточная стенка бактерий. Функции клеточной стенки. Строение клеточной стенки бактерии. Пептидогликан. Муреиновый мешок. Структура пептидогликана (муреина)
5. Грамотрицательные бактерии. Клеточная стенка грамотрицательных бактерий. Строение клеточной стенки грамотрицательных бактерий.
6. Грамположительные бактерии. Клеточная стенка грамположительных бактерий. Строение клеточной стенки грамположительных бактерий. Аутолизины бактерий. Сферопласты. Протопласты.
7. Цитоплазматическая мембрана ( ЦПМ ) бактерии. Состав цитоплазматической мембраны бактерий. Транспортные системы. Мезосомы. Периплазматическое пространство.
8. Цитоплазма бактерий. Бактериальный геном. Бактериальные рибосомы. Запасные гранулы бактерии.
9. Физиология бактерий. Питание бактерий. Тип питания бактерии. Голозои. Голофиты. Вода. Значимость воды для бактерий.
10. Усваиваемые бактериальной клеткой соединения. Пути поступления веществ в бактериальную клетку. Пассивный перенос. Диффузия.
Клеточная стенка бактерий. Функции клеточной стенки. Строение клеточной стенки бактерии. Пептидогликан. Муреиновый мешок. Структура пептидогликана (муреина)
Основные функции клеточной стенки следующие.
• Клеточная стенка защищает бактерии от внешних воздействий, придаёт им характерную форму, поддерживает постоянство внутренней среды и участвует в делении.
• Через клеточную стенку бактерий осуществляется транспорт питательных веществ и выделение метаболитов,
• На поверхности клеточной стенки располагаются рецепторы для бактериофагов, бактериоци-нов и различных химических веществ.
• Структура и состав элементов клеточной стенки определяет антигенную характеристику бактерий (по структуре О- и Vi-Аг).
• Клеточная стенка способна по-разному воспринимать красители; на этом основаны тинкториальные свойства бактерий.
• Нарушение синтеза компонентов клеточной стенки приводит к гибели бактерии или образованию 1-форм.
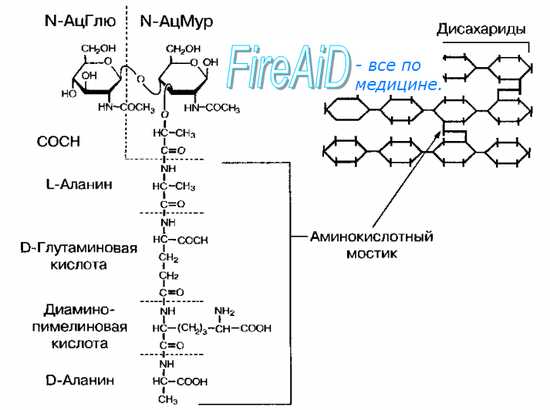
Рис. 4-3. Структура пептидогликана (муреина) Escherichia coll. Гетерополимерные цепочки, состоящие из чередующихся остатков N-ацитилглюкозамина (N-АцГлю) и N-ацетилмурамовой кислоты (N-АцМур), связаны между собой аминокислотными мостиками. Слева— детальное строение фрагмента пептидогликана. Справа — схематическое изображение структуры однослойного муреи-нового мешка.
Пептидогликан. Муреиновый мешок. Структура пептидогликана (муреина)
Опорный каркас клеточной стенки бактерий — пептидогликан (муреин) — гетерополимер, состоящий из повторяющихся дисахаридных групп, соединённых поперечными и боковыми цепочками (рис. 4-3). Остов молекулы пептидогликана — дисахарид. Его образуют N-ацетилглюкозамин и N-ацетилмурамовая кислота, соединённые через р-гликозидные связи. К молекуле N-ацетилмурамовой кислоты присоединяются олигопептиды, образующие боковые цепочки.
Связывание фрагментов пептидогликана заключается в образовании пептидной связи между терминальным остатком аминокислотного мостика (D-аланином) с предпоследним остатком примыкающего аминокислотного мостика (L-лизином или диаминопимелиновой кислотой в зависимости от вида бактерии). Боковые мостики образуют четыре аминокислоты, поперечные (вертикально связывающие слои пептидогликана) — пять аминокислот. В состав боковых мостиков также входят уникальные аминокислоты, присутствующие только у прокариотов, например диаминопимелиновая кислота (у большинства грамотрицательных бактерий) и D-изомеры глутаминовой кислоты и аланина. Эти соединения — «ахиллесова пята» бактерий, так как препараты, ингибирующие синтез компонентов клеточной стенки, обычно безвредны для растений и животных.
Пептидогликан лабилен к действию различных агентов. В частности, лизоцим гидролизует пептидогликан, расщепляя гликозидные связи между N-ацетилглюкозамином и N-ацетилмурамовой кислотой, то есть лизоцим действует как N-ацетилмурамидаза. Пептидазы расщепляют межпептидные связи, амидазы — связи между N-ацетилмурамовой кислотой и боковым пептидом (L-аланином). Лизостафин разрушает поперечные мостики пептидогликанов стафилококков. В-Лактамные антибиотики нарушают связывание боковых мостиков.
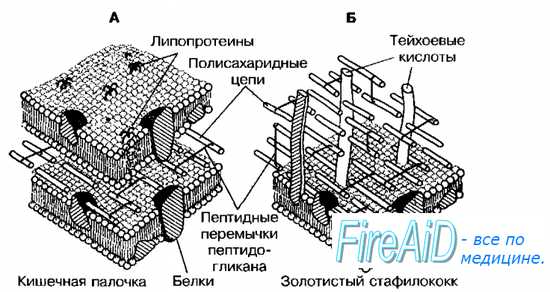
Рис. 4-4. Клеточная стенка грамотрицательных (А) и грамположительных (Б) бактерий.
Гетерополимерные пептидогликановые цепочки, соединённые пептидными связями, образуют гигантскую мешковидную макромолекулу («муреиновый мешок»), покрывающую всё тело бактерии. На поверхности «муреинового мешка» и в его толще располагаются различные вещества, характер и содержание которых лежит в основе разделения бактерий на грамположительные и грамотрицателъные (рис. 4-4).
– Также рекомендуем “Грамотрицательные бактерии. Клеточная стенка грамотрицательных бактерий. Строение клеточной стенки грамотрицательных бактерий.”
Источник
Кишечная палочка (син.: Bacterium coli commune, Escherichia coli) — грамотрицательная палочка семейства Enterobacteriaceae, рода Escherichia. Впервые выделена и описана в 1885 г. Т. Эшерихом. По систематике комитета общества американских бактериологов (Soc. Amer. Bact, committee, 1920) относится к роду Escherichia Castellani and Chalmers, включающему 22 вида.
Подкомитет по энтеробактериям (Enterobacteriaceae Subcommittee, 1962) отнес E. coli к группе Escherichia — Shigella, к-рая по классификации Юинга и Кауффманна представлена одноименным трибом, включающим два рода — Escherichia и Shigella. Род Escherichia по данной классификации и классификации определителя бактерий Берджи (Bergey’s Manuel of Determinative Bacteriology) включает только один вид — E. coli. Последний по антигенной структуре подразделяется на О- и ОК-серогруппы и на разнообразные биотипы: серологические, ферментативные, фаготипы, колицино- и колициногенотипы.
Одни из биотипов Кишечной палочки (так наз. банальные штаммы) являются комменсалами кишечного тракта людей и животных, другие относятся к паразитам, вызывающим заболевания.
Многие банальные штаммы Кишечной палочки обладают антагонистическими свойствами, препятствуя развитию в кишечнике патогенных энтеробактерий, дрожжеподобных грибков и других микробов. Некоторые штаммы Кишечной палочки синтезируют витамин В12 и другие факторы роста, покрывая в определенной мере потребности в них организма хозяина. Длительное применение антибиотиков широкого спектра действия приводит к гибели К. п. и развитию дисбактериоза (см.).
Кишечная палочка как постоянный обитатель кишечного тракта (см. Кишечник, микрофлора) выделяется с фекалиями в окружающую среду (почву, водоемы), где она не размножается, но сохраняет свою жизнеспособность примерно такой же срок, как и патогенные энтеробактерии (шигеллы, сальмонеллы). Поэтому она является санитарно-показательным микроорганизмом. Обнаружение К. п в исследуемых пробах указывает на их фекальное загрязнение (см. Коли-индекс, коли-титр), при к-ром наряду с К. п. могут встретиться возбудители кишечных инфекций.
Определенные биотипы К. п. могут явиться возбудителями острых кишечных инфекций и других заболеваний человека (см. Коли-инфекция). Их относят к энтеропатогенным К. п. (син. энтеропатогенные эшерихии). К. п., продуцирующие энтеротоксин, называют энтеротоксигенными. Дифференцировка этих бактерий производится по О- и OK-антигенам, что позволяет отнести их к определенным серогруппам и серотипам. Установлена этиологическую роль E. coli серогрупп 026, 055, 086, 0111, 0126 и др. при колиэнтеритах детей раннего возраста; серогрупп 025, 028 а, с, 032, 0124, 0144 и др. при дизентериеподобных заболеваниях взрослых и детей; серогрупп 01, 06, 015, 078, 0112 а, b,0148 при холероподобных заболеваниях людей. При инфекциях мочевыводящих путей встречаются Е. coli серогрупп 02, 06, 09, 018 и др., при аппендиците — 01, 02, 08, 015 и др., при холецистите — 01, 08, 011. При колибактериозе у животных выделены от телят Е. coli серогрупп 08, 09, 078, 0115 и др., от поросят — 08, 0138, 0141, 0149 и др. Иногда К. п. вызывают колисепсис, перитонит, эндотоксический (септический) шок (см. Аутоинтоксикация), пищевые токсикоинфекции.
Кишечная палочка используется как универсальная модель в общей и молекулярной генетике. Изучение огромного числа разнообразных мутантов одного из стандартных штаммов К. п.— Е. coli К-12 дало возможность составить генную карту и генный каталог бактериальной хромосомы (см. Бактерии).
Экология
Резервуаром Кишечной палочки в природе является человек, толстая кишка к-рого заселяется разными биотипами этого микроба с момента перехода ребенка на смешанное питание, примерно к концу первого года жизни. Количество К. п. в 1 г испражнений колеблется от нескольких миллионов до 1—3 млрд. особей. На протяжении жизни человека происходит многократная смена биотипов К. п. в кишечнике. Определенную роль в этом процессе играет режим питания, перенесенные инфекции, лечение химиопрепаратами, антибиотиками и другие факторы. В естественных условиях К. п. обитает также в кишечнике домашних животных, птиц, диких млекопитающих, рептилий, рыб и многих беспозвоночных.
Морфология

Электронограмма ультратонкого среза клетки E. coli: 1 — нуклеоид; 2 — цитоплазма; 3 — цитоплазматическая мембрана; 4 — клеточная стенка.
Клетки E. coli имеют форму палочек с закругленными концами длиной 1—2 мкм, толщиной 0,4—0,6 мкм (рис.).
Кишечная палочка грамотрицательна, ультраструктура сходна с другими грамотрицательными бактериями. Наряду с подвижными встречаются неподвижные формы.
Жгутики расположены перитрихиально. Спор не образуют. Встречаются штаммы с выраженной капсулой, у некоторых штаммов обнаружены реснички (пили).
Культуральные признаки
Культуры Кишечной палочки хорошо растут на обычных питательных средах при pH 7,2—7,4 и оптимальной t° 37°. К. п. является факультативным анаэробом (см. Анаэробы ). На плотных питательных средах образует гладкие, плоско-выпуклые, круглые, опалового цвета мутноватые, колонии. На среде Эндо колонии К. п. окрашены в красно-фиолетовый цвет с металлическим блеском. При росте на жидких средах К. п. дает помутнение и осадок.
Ферментативные свойства
К. п. продуцирует многочисленные сахаролитические ферменты, быстро ферментирует глюкозу и другие углеводы, чаще всего с кислото- и газообразованием. Почти все биотипы К. п. постоянно ферментируют маннит, арабинозу, мальтозу с образованием к-ты; св. 90% штаммов — лактозу, сорбит; непостоянно — сахарозу, раффинозу, рамнозу, ксилозу, дульцит, салицин; как правило, не ферментируют адонит и инозит. К. п. не утилизирует цитрат аммония, малонат натрия, не растет на среде с цианистым калием, не редуцирует нитраты в нитриты, не расщепляет мочевину, не разжижает желатину, большинство штаммов образует индол и не выделяет H2S. К. п. дает положительную реакцию с метилротом и отрицательную реакцию Фогеса — Проскауэра (см. Фогеса-Проскауэра реакция), не синтезирует ферменты цитохромоксидазу и фенилаланиндезаминазу, непостоянно декарбоксилирует лизин, орнитин и дегидролизует аргинин.
Антигены
Кишечная палочка содержит О-, К- и Н-антигены. О(соматические)-антигены у К. п., шигелл и сальмонелл имеют сходное хим. строение и связаны с липополисахаридом (ЛПС) клеточной стенки (см. Липополисахариды). Иммунохим, специфичность О-антигена определяется составом гексасахаридов в повторяющихся звеньях концевого участка полисахаридной цепи, к-рая другим концом связана через 2-кето-З-дезоксиоктонат (КДО) с липидом А (I). Так, структура ЛПС Е. coli 0111: В4 представлена следующим образом:
Обозначения: Кол — колитоза, Гл — глюкоза, Гал — галактоза, N-АцГл — N-ацетилглюкозамин, Геп – гептоза, х — неопределенный компонент.
Количество сахаров в одном и том же концевом звене, так же как и число детерминантных звеньев у разных серогрупп эшерихий, неодинаковое. Специфичность О-антигенов К. п. обычно определяется в реакции агглютинации (см.) с О- или ОВ-агглютинирующими колисыворотками на предметном стекле, реже другими методами.
К-антигенами обозначают поверхностные антигены, которые связаны с капсулой и с ЛПС К. п. Их подразделяют на А-, В- и L-антигены, отличающиеся друг от друга чувствительностью к высокой температуре и хим. агентам. Наиболее высокой устойчивостью к нагреванию (до 100° в течение 2,5 час.), спирту и 1 н. р-ру HCl обладает А-антиген, наименее устойчив L-антиген. У большинства К. п. К-антигены представляют собой кислые полисахариды, содержащие уроновые к-ты. Некоторые К-антигены (К 88) содержат только белок. К. п., имеющие К-антигены, не агглютинируются гомологичной О-коли-сывороткой. Эта особенность присуща живым культурам и утрачивается после их кипячения или автоклавирования. Наличие К-антигенов устанавливается также в реакции адсорбции агглютининов и при иммуноэлектрофоретическом исследовании (см. Иммуноэлектрофорез).
Н-жгутиковые, или флагеллярные, антигены присущи активно подвижным штаммам. Они связаны с белком-флагеллином и определяют типовую иммунохимическую специфичность К. п. Н-антигены термолабильны. Они полностью разрушаются при кипячении в течение 2,5 час.
У К. п. описано ок. 170 О-антигенов, 97 К-антигенов и 50 Н-антигенов. Штаммы К. п., снабженные ресничками, содержат белковые реснитчатые антигены. Их обнаруживают в реакции гемагглютинации (см.).
Св. 123 О-серологических групп эшерихий связаны друг с другом односторонними или двусторонними антигенными связями. Более чем у 56 серогрупп установлены антигенные связи с шигеллами и у 42 серогрупп с другими представителями семейства Enterobacteriaceae.
К. п. обозначаются по антигенным формулам: на первом месте ставится номер О-антигена, на втором — К-антигена, на третьем Н-антигена. Тип К-антигена указывается в скобках. Напр., К(А), К (В) или K(L). Номера О-, К- и Н-антигенов разделяются двоеточиями. По антигенным формулам устанавливается принадлежность исследованного штамма к определенной серол, группе [напр., Е. coli 026:К60 (B6)] и серол, типу [напр., Е. coli 026:К60 (В6):Н2]. Различия в рецепторах (факторном составе) О-антигенов обозначаются малыми буквами лат. алфавита. Напр., серогруппа 0111 :К58 (В4) разделена на 0111a,b:К58(В4) и 0111a,c:К58(В4).
Плазмиды
Кишечные палочки могут содержать различные плазмиды (см.): профаг, F (фактор фертильности), Col (колициногенный фактор), R (фактор резистентности), К88 (антигенный фактор), Ent (энтеротоксигенный фактор) и др. Профаги могут вызывать изменения отдельных признаков К. п., напр. О-антигена (см. Лизогения). F-плазмиды определяют половую полярность и вызывают образование половых ресничек (sex-pili). Col-плазмиды контролируют образование колицинов, подавляющих рост филогенетически родственных бактерий. R-плазмиды ответственны за резистентность К. п. к антибиотикам. Плазмида К88 контролирует синтез одноименного антигена и нитей, определяющих адгезивные свойства энтеропатогенных и энтеротоксигенных К. п.; плазмиды могут быть двух типов. Один из них определяет синтез термолабильного, другой — термолабильного и термостабильного энтеротоксина.
Патогенность и вирулентность
Кишечные палочки — комменсалы толстого кишечника — могут вызывать гнойно-воспалительные процессы в органах и тканях при резком снижении резистентности организма.
Вирулентность К. п. проявляется в адгезивности К. п., т. е. прилипании к ворсинкам эпителия кишки, размножении в просвете тонкой и толстой кишки, пенетрации в клетки эпителия и во внутриклеточном размножении, а также в подавлении фагоцитарной активности макрофагов и полиморфно-ядерных лейкоцитов (см. Вирулентность). К. п.— возбудители колиэнтеритов у детей раннего возраста и холероподобных заболеваний у взрослых — размножаются на поверхности эпителиальных клеток кишки. К. п. — возбудители дизентериеподобных заболеваний — проникают в клетки эпителия и размножаются в них так же, как шигеллы. Проявление вирулентности К. п. зависит от дозы бактерий, проникших в кишечник человека. Вирулентность К. п. определяется в тканевых культурах, опытах на лабораторных животных при воспроизведении экспериментального колиэнтерита, пневмонии и других процессов. Пенетрационная способность устанавливается кератоконъюнктивальной пробой Шереня путем введения культуры К. п. на конъюнктиву глаза морской свинки. К. п. образует эндотоксин, связанный с ЛПС, который является частью О-антигена. Биол, активность эндотоксина выражается неодинаково у равных серогрупп К. п. Токсические свойства эндотоксина определяются целой молекулой ЛПС, поскольку один липид или полисахарид малотоксичны. Эндотоксин поражает свертывающую систему крови, вызывает феномен Швартцмана (см. Швартцмана феномен) и другие явления, обладает пирогенными, адъювантными, протективными и митогенными свойствами. В небольших дозах стимулирует, а в больших угнетает фагоцитарную реакцию.
Многие серогруппы К. п. продуцируют энтеротоксины. Более постоянно их образует Е. coli серогрупп 06, 08, 015, 075, 078, 0148 и др. Полагают, что одни серогруппы К. п. вызывают холероподобную диарею у людей, другие только у животных (поросят, телят).
К. п. продуцируют два типа энтеротоксина. Термостабильный энтеротоксин инактивируется только после кипячения в течение 30 мин., медленно диализируется через целлофан, сохраняет активность при кислых значениях pH и после обработки трипсином и проназой. Молекулярный вес 10^3 — 10^4. Антигенными свойствами не обладает. Термолабильный энтеротоксин инактивируется после 30-минутного прогревания при 60°, pH 4,0—5,0, под действием проназы и не дуализируется. Мол. вес точно не установлен. Антигенная специфичность термолабильных энтеротоксинов, продуцируемых разными серогруппами К. п., и холерогена одинакова. Они стимулируют активность аденилциклазы и вызывают накопление циклического аденозинмонофосфата (цАМФ), что приводит к нарушению секреции и развитию острой диареи.
Для обнаружения энтеротоксигенных К. п. используют их способность вызывать расширение перевязанных участков тонкой кишки кролика и образование серозно-геморрагического экссудата. Другие методы основаны на способности термолабильного энтеротоксина активировать аденилциклазу и вызывать накопление в культуре ткани (яичников китайских хомячков, щитовидной железы свиньи) цАМФ, что приводит к индукции синтеза разных метаболитов и морфологическим изменениям клеток.
Резистентность
Устойчивость Кишечной палочки к воздействию внешних факторов — обычная для аспорогенных бактерий. Во внешней среде (воде, почве) она выживает в зависимости от конкретных условий в течение нескольких месяцев При нагревании во взвесях погибает при 55° через час, при 60° — через 15 мин., в 1% р-ре фенола — через 10 мин., в р-ре сулемы 1:4000 — через 2 мин. К. п. обладает избирательной чувствительностью к бриллиантовому зеленому и солям тетратионовой к-ты. На этом основано применение ряда селективных сред. Многие штаммы К. п. высокочувствительны к мономицину, канамицину, гентамицину.
Методы выделения и идентификации. Материалом для выделения К. п. являются объекты внешней среды (вода, почва, смывы с разных предметов), при заболеваниях — испражнения, рвотные массы, моча, дуоденальное содержимое или кровь, а также пищевые продукты, подозреваемые как источник заражения. Материал засевают на дифференциально-диагностические среды (см.) с последующим выделением чистой культуры К. п. Идентификацию последней проводят путем изучения морфологических, культуральных, биохимических и антигенных признаков. Заключительным этапом является определение биотипа Кишечной палочки, особенно серогруппы и серотипа.
См. также Escherichia.
Библиография: Борисов Л. Б. Энтеропатогенные кишечные палочки и их фаги, Л., 1976, библиогр.; Кауфман Ф. Семейство кишечных бактерий, пер. с англ., М., 1959, библиогр.; Методы санитарно-микробиологического исследования объектов окружающей среды, под ред. Г. И. Сидоренко, М., 1978; Минкевич И. Е. Бактерии группы кишечной палочки как санитарно-показательные микроорганизмы, Л., 1949; Острые кишечные инфекции, под ред. Т. В. Перадзе, с. 73, Л., 1973, библиогр.; Tабачник А. Л., Гиршович Е. С. и Темпер Р. М. Энтеротоксигенные Е. coli, Журн, микр., эпид, и иммун., № 3, с. 31, 1977, библиогр.; Bergey’s manual of determinative bacteriology, ed. by R. E. Buchanan a. N. E. Gibbons, Baltimore, 1975, bibliogr.; Cooke E. M. Escherichia coli and man, Edinburgh—L., 1974; Medearis D. N., Cammitta B. M. a. Heath E. C. Cell wall composition and virulence in Escherichia coli, J. exp. Med., v. 128, p. 399, 1968; Orskov I. a. o. Serology, chemistry and genetics of O and К antigens of Escherichia coli, Bact. Rev., v. 41, p. 667, 1977, bibliogr.
Л. Б. Борисов.
Источник