Лактозный оперон кишечной палочки
Регуляция активности генов у прокариот на примере лак-оперона
С-ва ген кода
Ген код – это принцип записи инф о последовательности аминокислот в полипептиде в виде последовательности нуклеотидов в молекуле Ирнк.
генетич код триплетен.
Триплеты в молекле иРНК наз кодонами, а комплементарные им тирплеты в молекуле тРНК- антиколонами.
свойства ген кода
1 . Триплетность.одну аминокислоту кодируют три рядом расположенных нуклеотида.
2. Неперекрываемость.каждый нуклеотид входит в состав только одного кодона.
3.Вырожденность (избыточность).один смысловой элемент (аминокислота) шифруется несколькими кодонами.
4. Специфичность (однозначность).каждый отдельный кодон кодирует только один аминокислотный остаток в молекуле полипептида.
5. Непрерывность.каждый нуклеотид принадлежит какому либо триплету т.е между кодонами иРНК нет нуклеотидов, не входящих в последовательность кодонов данного гена.
6. Коллинеарность.кодоны нуклеотдиных кислот и соответствующие им аминокислоты полипептидов расположены в одинаковом линейном порядке.
7. Однонаправленность. Считавание кода начинается с определяемой кодоном – инициатором точки и идёт в одном направлении в пределах данного гена от 5’концу к 3’концу.
8. Универсальность.ген код одинаков для всез организмов.
активность генов проявляется на уровне определяемых ими фенотипических эффектах. Мерой активности генов служит функциональная активность белков, контролируемых этими генами.
В механизме регуляции активности генов прокариот большую роль играют особые гены-регуляторы, контролирующие синтез регуляторных белков.
Такие белки, соединяесь с последовательностями промоторов реагируемых генов, способны подавлять или активировать их транскрипцию.
Регуляторные белки, подавляющие транскрипцию структурных генов, наз репрессорами.
Последовательности нуклеотидов регуляторных генов, с которыми взаимодействуют белки репрессоры, получили название операторов.
Регуляция, связанная с подавлением транскрипции, наз негативной.
Регуляторные белки, активирующие транскрипции. структурных генов, наз активаторами. Регуляция, связанная с активацией транскрипции получила наз – позитивной.
К негенетиским факторам регуляции экспрессии генов, или эффектрорам, относятся в-ва небелковой природы. Взаимодействуя с регуляторными бедками, они изменяют их биологическую активность. Различают 2 вида эффекторов: индукторы – «включающие» транскрипцию и корепрессоры, «выкл её».
Лактозный оперон E coil включ след элементы: 3 гена, кодирующих белки ферменты: B-галактозидазу, пермеазу и трансацетилазу, участвующие в метаболизме лактозы и транспорте её в клетку, и регуляторной области. Регуляторная область, в свою очередь, сост из промотора, оператора– последовательности нуклеотидов для связ белка репрессора, а также последовательности нуклеотидов для связ белка активатора. Активность генов контролир регуляторным геном Lac1.
При выращивании E.coli на среде, содержащей только глюкозу геи- регулятор лак-оперон синтезирует активный белок-репрсссор, который, взаимодействуя с оператором, «выключает» транскрипцию структурных генов, кодирующих ферменты, участвующие в метаболизме и транспорте лактозы в клетку.
Если клетки E.coli перенести на среду, содержащую только лактозу, то проникая внутрь клеток небольшая часть ее превращается в аллолактозу, которая связываясь с белком -репрессором, инактивируст его. В результате РНК-полимераза осуществляет транскрипцию полицистронной мРНК для синтеза всех ферментов, необходимых для транспорта и метаболизма лактозы.
В данном случает осуществляется негативная регуляция генов оперона. При этом аллолактоза служит ИНДУКАТОРОМ генов лак-оперона, кодирующего белки, участвующие в транспорте и метаболизме лактозы.
При культивировании кишечной палочки на среде, содержащей как лактозу, так и глюкозу клетки Е coli, используют для гликолиза в основном глюкозу. Указанная особенность метаболизма обусловливается наличием у Е coli механизма положительной регуляции активности генов lac оперона.
50. общая схема регуляции генов у эукариот
1) осуществление транскрипции эукриотических генов возможно лишь при декомпактизации хроматина: 2) регуляция активности генов у эукариот осуществляется на всех уровнях реализации наследственной информации: на уровне транскрипции, РНК -процессннта альтернативный сплайсинг), транспорта зрелой мРНК из ядра в цитоплазму, трансляции и посттрансляционных преобразований белков ( химическая модификация и разрушение функционально активного полипептида) 3)активность каждою структурного гена контролируется многими генами-регуляторами, а эффекторами часто служат гормоны.
Источник
Схема негативной индукции Жакоба и Моно
Lac-оперон E. coli содержит 3 гена, отвечающие за образование белков, участвующих в переносе в клетку дисахарида лактозы и в ее расщеплении.
Z – b- галактозидаза (расщепляет лактозу на глюкозу и галактозу).
Y- b- галактозидпермеаза (переносит лактозу через мембрану клетки).
А – тиогалактозидтрансацетилаза (ацетилирует галактозу).
В отсутствие в клетке лактозы lac- оперон выключен. Активный белок – репрессор, кодируемый в моноцистронном опероне (LacI) , не имеющем оператора, связан с оператором lac-оперона. Поскольку оператор перекрывается с промотором, даже посадка РНК-полимеразы на промотор невозможна.
Как только некоторое количество лактозы попадает в клетку, две молекулы субстрата (лактозы) взаимодействуют с белком – репрессором, изменяют его конформацию – и он теряеет сродство к оператору.
Тут же начинается транскрипция lac-оперона и трансляция образующейся mРНК; три синтезируемых белка участвуют в утилизации лактозы.
Когда вся лактоза переработана, очередная порция репрессора, свободного от лактозы, выключает lac-оперон.
Внимание!
Если вам нужна помощь в написании работы, то рекомендуем обратиться к
профессионалам. Более 70 000 авторов готовы помочь вам прямо сейчас. Бесплатные
корректировки и доработки. Узнайте стоимость своей работы.
Эта схема называется так потому, что контролирующим транскрипцию фактором является негативный фактор, “выключатель” – белок – репрессор. Индукция (включение) происходит при потере сродства белка – репрессора к оператору.
Существует и позитивная регуляция работы lac-оперона E. coli.
Схема позитивной индукции
Аra-оперон E. сoli.
В нем 3 цистрона, которые кодируют ферменты, расщепляющие сахар арабинозу. В норме оперон закрыт. Белок – репрессор связан с оператором. |
|
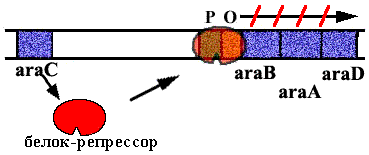
Когда в клетку попадает арабиноза, она взаимодействует с белком – репрессором. Белок – репрессор меняет конформацию и превращается из репрессора в активатор, взаимодейсивующий с промотором и облегчающий посадку РНК-полимеразы на промотор.
Эта схема регуляции называется позитивной индукцией, поскольку контролирующий элемент – белок – активатор “включает” работу оперона.
(другой вариант) Lac-оперон состоит из кодирующей области, представленной тремя
структурными генами, ответственными за синтез ферментов: β-галактозидазы, β-галактозидпермеазы и β-галактозидтрансацетилазы; а также из промоторно-операторной области. Оператор представляет собой небольшой участок ДНК, граничащий с первым структурным геном. С оператором может связываться белок-репрессор, блокируя инициацию транскрипции. Промотор – это небольшой участок ДНК перед оператором. Он служит местом связывания ДНК-зависимой РНК-полимеразы и от него начинается транскрипция ДНК. Оператор и промотор в некоторой степени перекрываются таким образом, что когда репрессор связан с ДНК в области оператора, то РНК-полимераза не может связаться с промотором и транскрипция структурных генов не происходит. Следовательно, промотор, оператор и структурные гены образуют оперон. Транскрипционная активность входящих в оперон генов регулируется специальным геном-регулятором, или регуляторным геном (ген R),который может располагаться рядом со структурными генами или на некотором расстоянии от них. Ген R кодирует синтез специфического белка-репрессора. Репрессор – аллостерический белок, имеющий два центра связывания: один центр узнает оператор, другой – взаимодействует с эффектором или индуктором. Для Lac-оперона индуктором является лактоза, которая связывается с репрессором, переводит его в неактивную форму, в результате чего репрессор отсоединяется от оператора.
Получить выполненную работу или консультацию специалиста по вашему
учебному проекту
Узнать стоимость
Источник
Оперон, обеспечивающий
у E.coliспособность к сбраживанию
молочного сахара – лактозы, состоит из
промотора, оператора и трех структурных
генов. Ген lac Z кодирует фермент-
генгалактозидазу, катализирующую
гидролиз лактозы до глюкозы и галактозы;
ген lac Y-галактозидпермеазу, обеспечивающую
транспорт различных сахаров, включая
лактозу, мелибиозу и рафинозу, в клетку;
ген lac А –тиогалактозидтрансацетилазу,
роль которой обычно в утилизации лактозы
не ясна. Все три белка обычно присутствуют
в клеткахЕ.coliв следовых количествах.
Однако при выращивании бактерий на
среде, в которой единственным источником
углерода и энергии служит лактоза,
количество указанных ферментов
увеличивается в 1000 раз.
Ген–регулятор
лактозного (lac) оперона, обозначаемый
lac 1, кодирует белок–репрессор. В активной
форме это тетрамер, образованный четырьмя
копиями продукта гена lac 1 – полипептидами,
состоящими из 360 аминокислот. Клетки с
мутациями в гене lac 1 конститутивны по
синтезу ферментов, кодируемых генами
lac Z, Y и A.
Конститутивный
синтез продуктов этих генов возможен
не только в случае lac 1 -мутаций в
гене репрессоре, но и в случае мутаций
в операторе, обозначаемых О![]() .
.
Такие мутации всегда цис–доминантны,
поскольку, в отличие от гена–репрессора,
оператор может влиять на возможность
транскрипции структурных генов только
тогда, когда он находится непосредственно
рядом с промотором. Если в клетке
находится индуктор, он конкурирует с
оператором за молекулы репрессора,
причем репрессор в первую очередь
связывается с индуктором.
В качестве индукторов
могут служить различные соединения.
Лактоза представляет собой индуктор и
одновременно субстрат. В нормальных
клетках даже в отсутствие индуктора
остаточная активность пермеазы и –галактозидазы обеспечивают возможность
проникновения в клетку минимального
количества лактозы, которая в результате
реакции, катализируемой–галактозидазой, переходит в аллолактозу.
Последняя связывается с репрессором,
обусловливая его отсоединение от
оператора, что, в свою очередь, открывает
путь РНК–полимеразе для связывания с
промотором и транскрипции генов lac Z, Y
и A. К соединениям, являющимся только
индукторами, но не субстратами, относятся
изопропил–D–тиогалактопиранозид
(ИПТГ) и тиометил––D–галактопиранозид
(ТМГ), часто используемые для исследования
регуляции lac–оперона.
Мутации в промоторе
в отличие от мутаций в гене–репрессоре
и в операторе не влияют на индуцируемость
оперона, однако они регулируют уровень
его экспрессии, изменяя эффективность
присоединения РНК–полимеразы, и тем
самым частоту инициации транскрипции
lac–оперона.
Наряду с негативной
системой регуляции, lac–оперон
контролируется и с помощью позитивно
действующих элементов. Их обнаружение
связано с исследованием феномена Ж.
Моно диаусией, суть которого состоит
в том, что утилизация лактозы начнется
лишь после того, как будет использована
вся имеющаяся в среде глюкоза. Этот
феномен, как установили Б. Магазаник с
соавторами, – одно из проявленийкатаболитной репрессии или глюкозного
эффекта, известного еще с 40-х годов и
выражающегося в неспособностиE.coli,
катаболизироватьразличные
углеводы (лактозу, арабинозу, галактозу
и др.) в присутствии глюкозы, как более
эффективного источника энергии.
Расшифровать
механизм глюкозного эффекта сумели Р.
Перлман и А. Пастан, обнаружившие, что
транскрипция lac–оперона контролируется
двумя элементами: небольшой
молекулой–эффектором, циклическим
аденозинмонофосфатом (цАМФ) и
белком–активатором САР (от первых букв
англ. Catabolite fctivator protein– белок-активатор
катаболизма), называемым также
белком–рецептором цАМФ. У эукариот
цАМФ является медиатором действия
гормонов. Оказалось, что добавление
цАМФ к растущим в среде с глюкозой
клеткам E .coliхотя и замедляет скорость
их роста, но снимает катаболическую
репрессию, обусловливая тем самым
возможность экспрессии лактозного
оперона в условиях одновременного
присутствия в среде лактозы и глюкозы.
Позднее была показана обратная зависимость
между содержанием в клетке цАМФ и
глюкозы: глюкоза подавляет активность
фермента, синтезирующего цАМФ из АТФ.
Этот фермент, названныйаденилатциклазой,
кодируется геномсуа.
В структуре
промотора lac–оперона выявлено два сайта
связывания. Один из них взаимодействует
с РНК – полимеразой, другой – с комплексом
САР–цАМФ. Присоединение комплекса
САР–цАМФ к своему сайту на промоторе
– условие индукции оперона. Следовательно,
этот комплекс позитивно контролирует
транскрипцию lac–оперона. Белок САР
состоит из двух идентичных субъединиц
с общей М![]() около 45000, кодируемых геном САР, илиCRP.
около 45000, кодируемых геном САР, илиCRP.
Мутации в генесарнарушают участок
связывания белка с цАМФ, либо расширяют
спектр кофакторов, объединение с которыми
обеспечивает индукцию ферментов
lac–оперона. У некоторых мутантов в генесартаким кофактором наряду с цАМФ
может служить и цГМФ.
В норме, то есть в
присутствии глюкозы и в отсутствии
цАМФ, белок САР не может объединяться
с промотором lac–оперона. В свою очередь,
РНК–полимераза не способна эффективно
связываться с этим промотором, если к
нему не присоединен комплекс САР–цАМФ.
Некоторые мутации в промоторе обусловливают
независимость экспрессии lac – оперона
от глюкозного эффекта, снижая сродство
промотора к комплексу САР–цАМФ.
Таким образом,
транскрипция lac–оперона на самом деле
находится под двойным – негативным и
позитивным– контролем. Комплекс САР–цАМФ
позволяет РНК–полимеразе присоединиться
к матричной ДНК до начала транскрипции.
Репрессор – продукт гена
lac 1–препятствует инициации синтеза
иРНК.
В настоящее время
расшифрована полная нуклеотидная
последовательность регуляторной области
lac–оперона, включающая промотор и
оператор. Более того, ДЖ. Шапиро и ДЖ.
Беквит с соавторами (1969) сумели выделить
чистую ДНК этого оперона, включающую
фрагмент гена lac1, полностью промоторную
и операторную последовательности, ген
lac Z , а также фрагмент гена lac Y. Выяснение
структурной организации оператора
lac–оперона показало, что существенную
роль во взаимодействиях мультимерных
белков типа lac–репрессора или
РНК–полимеразы с ДНК играют симметричные
структуры – палиндромы. Оператор
lac–оперона состоит из 26 п.н., из которых
14 представляют собой палиндром: в
различных цепях они читаются одинаково,
но в противоположных направлениях.
Палиндром обнаружен и в участке промотора,
связывающемся с комплексом САР–цАМФ.
Соседние файлы в предмете [НЕСОРТИРОВАННОЕ]
- #
- #
- #
- #
- #
- #
- #
07.03.201643.04 Mб23Мондор Г. – Неотложная диагностика. Живот (2 тома)Том 2.djvu
- #
- #
- #
- #
Источник
Схема строения и работы оперона
Регуляция у прокариот
Транскрипция хорошо регулируется
Внешний вид вторичной структуры тРНК после процессинга
Строение модифицированных уридиловых нуклеотидов
Процессинг предшественника транспортной РНК
1. Модификация нуклеотидов в молекуле путем дезаминирования, метилирования, восстановления.
Например, образование псевдоуридина и дигидроуридина.
2. Формирование антикодоновой петли происходит путем сплайсинга (вернуться вверх) и удаления интрона в средней части пре-тРНК.
3. Формирование на 3′-конце последовательности ЦЦА. Для этого у одних пре-тРНК с 3′-конца удаляются лишние нуклеотиды до “обнажения” триплета ЦЦА, у других идет присоединение этой последовательности.
Так как транскрипция связывает ядро– “мозг” клетки, ее “банк знаний” и белки, “рабочих лошадок” клетки, то от качества и активности транскрипции зависит объем синтеза тех или иных белков, жизнедеятельность клетки, ее способность адаптироваться к окружающей обстановке.
У прокариот и эукариот регуляция транскрипции происходит, естественно, по-разному, хотя некоторые моменты похожи.
Регуляция биосинтеза белка у прокариот осуществляется на уровне изменения скорости синтеза мРНК. В настоящее время принята теория оперона, сформулированная Франсуа Жакобом и Жаком Моно. В основе теории лежат следующие понятия:
· оперон– группа тесно связанных между собой генов, которые программируют образованиеструктурных белков и ферментовв клетке,
· конституитивные ферменты – те, которые присутствуют в клетках всегда, независимо от ее активности и условий,
· индуцибельные ферменты – те, которые программируются опероном и синтезируются при необходимости,
· ген-регулятор – ген, регулирующий работу оперона, но не входящий в его состав. Он синтезирует белок-регулятор (чаще называемый белок-репрессор), который может быть в активной или неактивной форме,
· ген-оператор – участок ДНК, способный связываться с белком-регулятором, и “решающий” нужно работать РНК-полимеразе или нет.
Предложены две схемы регуляции скорости транскрипции: по механизму индукции (лактозный оперон) и помеханизму репресии (триптофановый оперон).
Лактозный оперон в целом отвечает за катаболизм лактозы.
При изучении E.coli было замечено, что в клетке может быть две взаимоисключающие ситуации:
· активность одного из ферментов катаболизма лактозы низка, если в среде имеетсяглюкоза.
· активность этого фермента резко повышаетсяв обратной ситуации, т.е. при отсутствии глюкозыи приналичии лактозы.
На основании наблюдений была предложена схема регуляции оперона по механизму индукции:
1. В отсутствие лактозы активный белок-репрессор связывается с оператором и блокирует синтез мРНК, кодирующей ферменты катаболизма лактозы. В результате эти ферменты не образуются.
2. Если глюкозы нет, алактоза есть, то последняя связывается с белком-репрессором и ингибирует его, не давая связаться с геном-оператором. Это позволяет РНК-полимеразе считывать информацию, отвечающую за синтез ферментов катаболизма лактозы, и синтезировать мРНК.
Таким образом, лактоза является индукторомтранскрипции.
Источник