Переваривание нуклеиновых кислот в желудочно кишечном тракте
РАЗДЕЛ 10. ОБМЕН НУКЛЕОТИДОВ
Рибонуклеозид- и дезоксирибонуклеозидфосфаты — существеннейшие компоненты клеток.
• Нуклеозидтрифосфаты (НТФ) используются в качестве субстратов синтеза ДНК и РНК, без которых невозможны образование белков и клеточная пролиферация.
• Природа выбрала цикл АДФ-АТФ в качестве универсального механизма трансформации энергии окисления в энергию биосинтетических процессов. В некоторых биологических процессах и другие НТФ используются в качестве источника энергии.
• Производные нуклеотидов служат донорами активных субстратов в синтезе гомо- и гетерополисахаридов, липидов и белков. Например, УДФ-глюкоза, УДФ-галактоза, ГДФ-манноза, УДФ- N-ацетилглюкозамин или ЦМФ-ацетилнейраминовая кислота принимают участие в синтезе гликогена и гликозаминогликанов; ЦЦФ-холин — в синтезе фосфолипидов.
• УДФ-глюкуроновая кислота, ФАФС, S-аденозилметионин — наиболее частые участники универсальной системы детоксикации, обеспечивающей последующее выведение ксенобиотиков (чужеродных веществ) и некоторых собственных метаболитов из организма.
• АМФ входит в состав коферментов дегидрогеназ (NAD+, NADP+, FAD, FMN) и ацилирования (КоА).
• С помощью циклических форм нуклеотидов (цАМФ, цГМФ) осуществляется передача в клетку сигналов гормонов, факторов роста, нейромедиаторов и некоторых других регуляторных молекул.
Практически все клетки организма способны к синтезу нуклеотидов (исключение составляют некоторые клетки крови). Другим источником этих молекул могут быть нуклеиновые кислоты собственных тканей и пищи, однако эти источники имеют лишь второстепенное, вспомогательное значение.
Со строением нуклеотидов и номенклатурой мы знакомились в разделе 4, в данном разделе предстоит рассмотреть метаболизм этих молекул в организме.
I. Переваривание нуклеиновых кислот пищи в желудочно-кишечном тракте
Пищевые нуклеопротеины, попадая в организм человека, в желудке отщепляют белковый компонент и денатурируют под действием НСl желудочного сока (рис. 10-1). Далее полинуклеотидная часть этих молекул гидролизуется в кишечнике до мононуклеотидов.
Рис. 10-1. Переваривание нуклеиновых кислот пищи.

В расщеплении нуклеиновых кислот принимают участие ДНК-азы и РНК-азы панкреатического сока, которые, будучи эндонуклеазами, гидролизуют макромолекулы до олигонуклеотидов. Последние под действием фосфодиэстераз панкреатической железы расщепляются до смеси 3′- и 5′-мононуклеотидов. Нуклеотидазы и неспецифические фосфатазы гидролитически отщепляют фосфатный остаток нуклеотидов и превращают их в нуклеозиды, которые либо всасываются клетками тонкого кишечника, либо расщепляются нуклеозидфосфорилазами кишечника с образованием рибозо- или дезоксирибозо-1-фосфата, пуриновых и пиримидиновых оснований.
Пищевые пурины и пиримидины не являются незаменимыми пищевыми факторами и очень мало используются для синтеза нуклеиновых кислот тканей. В энтероцитах обнаружена высокая активность ксантиноксидазы — фермента, который большую часть пуринов, поступающих в клетки, превращает в мочевую кислоту, удаляющуюся с мочой. Пиримидиновые основания, не успевшие поступить в энтероциты, под действием микрофлоры кишечника расщепляются до NH3, СО2, β-аланина и β-аминоизобутирата.
В различных клетках организма синтезируется до 90% пуриновых и пиримидиновых нуклеотидов из простых предшественников de novo. Введённые в кровь азотистые основания и нуклеозиды, а также основания и нуклеозиды, образующиеся в результате внутриклеточного разрушения нуклеиновых кислот, в небольшом количестве могут использоваться для повторного синтеза нуклеотидов по так называемым «запасным» путям.
Источник
Пищевые нуклеопротеины,
попадая в организм человека, в желудке
отщепляют белковый компонент и
денатурируют под действием НС1 желудочного
сок. Далее полинуклеотидная часть этих
молекул гидролизуется в кишечнике до
мононуклеотидов.
В расщеплении нуклеиновых
кислот принимают участие ДНК-азы
и РНК-азы панкреатического
сока, которые, будучи эндонуклеазами,
гидролизуют макромолекулы до
олигонуклеотидов. Последние под действием
фосфодиэстераз панкреатической железы
расщепляются до смеси 3′- и 5′-мононуклеотидов.
Нуклеотидазы и
неспецифические фосфатазы
гидролитически отщепляют фосфатный
остаток нуклеотидов и превращают их в
нуклеозиды, которые либо всасываются
клетками тонкого кишечника, либо
расщепляются нуклеозидфосфорилазами
кишечника с образованием рибозо- или
дезоксирибозо-1-фосфата, пуриновых и
пиримидиновых оснований.
Пищевые пурины и пиримидины
не являются незаменимыми пищевыми
факторами и очень мало используются
для синтеза нуклеиновых кислот тканей.
В энтероцитах обнаружена высокая
активность ксантиноксидазы – фермента,
который большую часть пуринов, поступающих
в клетки, превращает в мочевую кислоту,
удаляющуюся с мочой. Пиримидиновые
основания, не успевшие поступить в
энтероциты, под действием микрофлоры
кишечника расщепляются до NH3,
CO2,
β-аланина и β-аминоизобутирата.
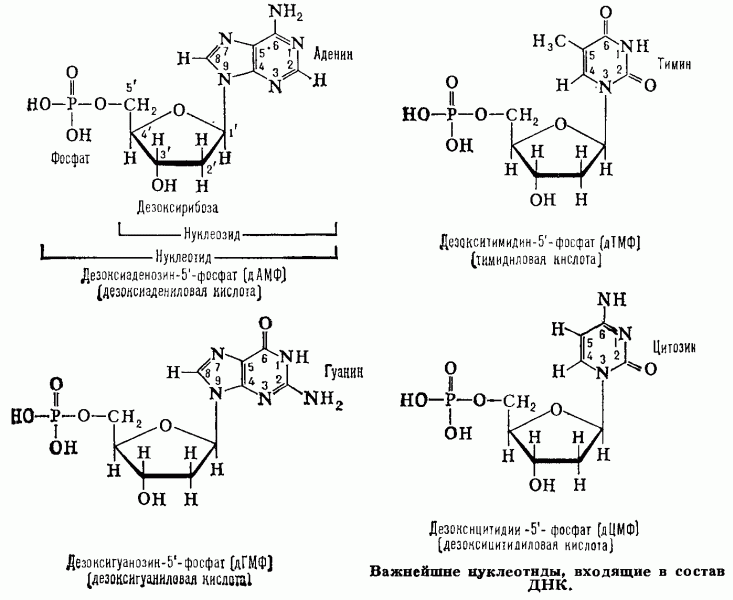
Вопрос № 2.Структура нуклеотида
В
каждом нуклеотиде можно выделить 3
части:
1.
Углевод – это пятичленный сахар-пентоза
(рибоза или дезоксирибоза).
2.
Фосфорный остаток (фосфат) – это остаток
фосфорной кислоты.
3.
Азотистое основание – это соединение, в
котором много атомов азота. В нуклеиновых
кислотах используется всего 5 видов
азотистых оснований : Аденин, Тимин,
Гуанин, Цитозин, Урацил. В ДНК – 4 вида:
Аденин, Тимин, Гуанин, Цитозин. В РНК
– тоже 4 вида: Аденин, Урацил, Гуанин,
Цитозин, Легко заметить, что в РНК
происходит замещение Тимина на Урацил
по сравнению с ДНК.
Общая
структурная формула пентозы (рибозы
или дезоксирибозы), молекулы которой
образуют “скелет” нуклеиновых
кислот:
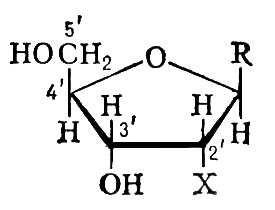
Если
Х заменить на Н (Х = Н) – то получаются
дезоксирибонуклеозиды; если Х заменить
на ОН (Х = ОН) – то получаются рибонуклеозиды.
Если вместо R подставить азотистое
основание (пуриновое или пиримидиновое)
– то получится конкретный нуклеотид.
Важно обратить внимание на
те положения атомов углерода в
пентозе, которые обозначены как 3′ и
5′. Нумерация атомов углерода начинается
от атома кислорода вверху и идёт по
часовой стрелке. Последним получается
атом углерода (5′), который располагается
за пределами пентозного кольца и
образует, можно сказать, “хвостик”
у пентозы. Так вот, при наращивании
цепочки из нуклеотидов фермент может
присоединить новый нуклеотид только к
углероду 3′ и ни к какому другому. Поэтому
5′-конец нуклеотидной цепочки никогда
не сможет иметь продолжения, удлинняться
может только 3′-конец.
Азотистые основания
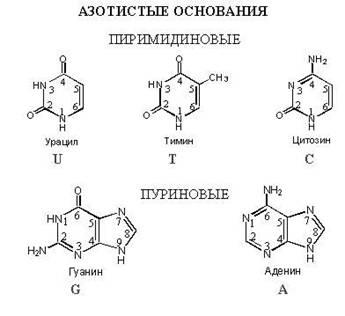
Нуклеотиды
Наращивание нуклеотидной цепочки днк
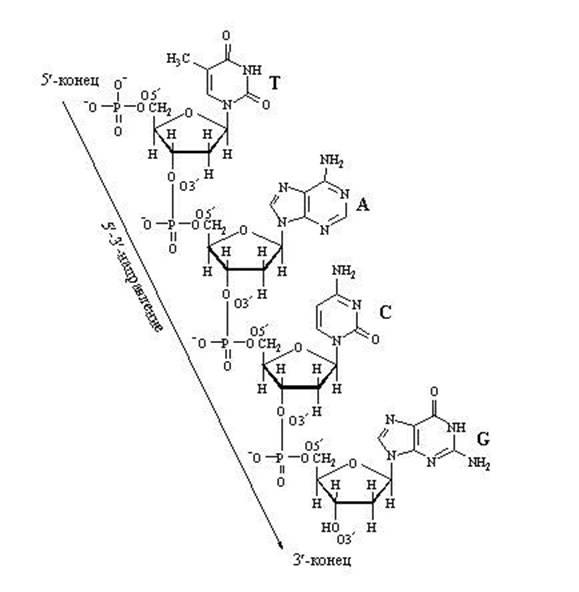
Наращивание цепочки за
счёт “продольных” связей между
нуклеотидами может идти только в одном
направлении: от 5′ ⇒
к 3′, т.к. новый нуклеотид можно присоединить
только к 3′-концу цепочки, но не к
5′-концу.
Соседние файлы в предмете [НЕСОРТИРОВАННОЕ]
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
Источник
С пищей в сутки в организм поступает около 1 г нуклеиновых кислот.
Переваривание нуклеиновых кислот происходит в тонком кишечнике. Сначала, поступившие с пищей нуклеиновые кислоты под действием ферментов панкреатического сока – нуклеаз – превращаются в мононуклеотиды. Затем уже под влиянием ферментов тонкого кишечника от мононуклеотидов отщепляется фосфорная кислота, и образуются нуклеозиды. Часть нуклеозидов расщепляется затем на азотистое основание и углевод.
Продукты переваривания нуклеиновых кислот поступают в кровь, а затем в печень и другие органы.
В клетках организмов обмен РНК протекает значительно более интенсивно, чем обмен ДНК. В конечном итоге нуклеиновые кислоты расщепляются на азотистые основания, углеводы и фосфорную кислоту.
Далее пуриновыеазотистые основания в процессе катаболизма теряют аминогруппу в виде аммиака, окисляются и превращаются в мочевую кислоту.
Пиримидиновые основания подвергаются более глубокому расщеплению до углекислого газа, воды и аммиака.
Углеводывовлекаются в ГМФ-путь распада и превращаются в глюкозу.
Фосфорная кислотараспаду не подвергается. Она используется в реакциях фосфорилирования и фосфолиза или при избытке выделяется из организма с мочой.
Синтез нуклеотидов.
Все клетки организма способны синтезировать необходимые нуклеиновые кислоты и не нуждаются в наличии в пище готовых нуклеиновых кислот или их составных частей. Поэтому содержание готовых нуклеиновых кислот в пище для организма принципиального значения не имеет, хотя продукты их распада могут частично использоваться организмом.
Синтез пуриновых и пиримидиновых нуклеотидов происходит на основе рибозо-5-фосфата. из глюкозы при её распаде по ГМФ-пути. Свободные азотистые основания обычно для этого синтеза не используются.
При синтезе пуриновых нуклеотидов к рибозо-5-фосфату присоединяются атомы углерода и азота, из которых образуется пуриновое кольцо. Источниками этих атомов являются аминокислоты глицин, глутамин, аспарагиновая кислота.Часть атомов углерода поставляется коферментами, содержащими в своём составе фолиевую кислоту и биотин. Промежуточным продуктом синтеза пуриновых нуклеотидов является инозиновая кислота. Далее из инозиновой кислоты образуются пуриновые нуклеотиды.
Синтезу пиримидиновых нуклеотидов предшествует образование необычного азотистого основания оротовой кислоты,содержащей пиримидиновое кольцо. Синтезируется оротовая кислота из аммиака и аспарагиновой кислоты. Оротовая кислота присоединяется к рибозо-5-фосфату и возникает пиримидиновый нуклеотид оротидинмонофосфат.Далее оротовая кислота в составе этого нуклеотида преобразуется в обычные азотистые основания, в результате чего появляются пиримидиновые нуклеотиды.
В связи с высокой важностью оротовой кислоты в спортивной практике в качестве пищевой добавки используется её соль оротат калия.
Дезоксирибонуклеотидыобразуются из соответствующих рибонуклеотидов путём восстановления входящей в них рибозы в дезоксирибозу.
Синтез нуклеиновых кислот.
Для синтеза нуклеиновых кислот используются мононуклеотиды обязательно в трифосфатной форме. Такие нуклеотиды содержат в своей молекуле три остатка фосфорной кислоты и обладают повышенным запасом энергии. Переход нуклеотидов в трифосфатную форму осуществляется путем взаимодействия с АТФ. Для синтеза РНК используются АТФ, ГТФ, УТФ, ЦТФ. А для синтеза ДНК, соответственно, дАТФ, дГТФ, дТТФ, дЦТФ.
Процесс репликации или редупликацииДНК иначе можно назвать удвоением.Он происходит перед делением клетки. Осуществляет его специальный фермент ДНК полимераза. Этот фермент разделяет две цепи двойной спирали и достраивает к каждой из них комплементарную ей цепь. Таким образом, из одной молекулы образуется две одинаковые дочерние молекулы, причем обе цепи ДНК служат матрицами для дочерних цепей. По мере присоединения к матрице нуклеотиды связываются в полинуклеотидные нити, которые сразу же закручиваются в двойную спираль. Биологический смысл репликации состоит в том, что из одной молекулы ДНК появляются две ее полные копии. Процесс этот идет с очень высокой точностью – ошибки крайне редки.
Процесс синтеза РНК называется транскрипцией. Процесс образования белков на матрицах информационной РНК называется трансляцией.
Транскрипцию осуществляет фермент РНК-полимераза. Этот фермент соединяет между собой рибонуклеотиды, составляющие остов молекулы РНК. Делает это фермент на основании считывания последовательности молекулы ДНК и, достраивая комплементарные ей последовательности. Показано, что в этом процессе только одна из двух цепей ДНК играет роль матрицы. Бывают, правда, и исключения – это ДНК некоторых вирусов. В процессе транскрипции участвует только ограниченный участок ДНК. Именно этот участок ДНК и понимают в молекулярной биологии, как ген.
ОБМЕН БЕЛКОВ.
1. Переваривание и всасывание белков.
2. Катаболизм белков.
3. Синтез белков.
4. Метаболизм аминокислот.
5. Азотистый баланс. Обезвреживание аммиака.
Источник
Пищевые нуклеопротеины, попадая в организм человека, в желудке отщепляют белковый компонент и денатурируют под действием НС1 желудочного сок. Далее полинуклеотидная часть этих молекул гидролизуется в кишечнике до мононуклеотидов.
В расщеплении нуклеиновых кислот принимают участие ДНК-азы и РНК-азы панкреатического сока, которые, будучи эндонуклеазами, гидролизуют макромолекулы до олигонуклеотидов. Последние под действием фосфодиэстераз панкреатической железы расщепляются до смеси 3′- и 5′-мононуклеотидов. Нуклеотидазы и неспецифические фосфатазы гидролитически отщепляют фосфатный остаток нуклеотидов и превращают их в нуклеозиды, которые либо всасываются клетками тонкого кишечника, либо расщепляются нуклеозидфосфорилазами кишечника с образованием рибозо- или дезоксирибозо-1-фосфата, пуриновых и пиримидиновых оснований.
Пищевые пурины и пиримидины не являются незаменимыми пищевыми факторами и очень мало используются для синтеза нуклеиновых кислот тканей. В энтероцитах обнаружена высокая активность ксантиноксидазы – фермента, который большую часть пуринов, поступающих в клетки, превращает в мочевую кислоту, удаляющуюся с мочой. Пиримидиновые основания, не успевшие поступить в энтероциты, под действием микрофлоры кишечника расщепляются до NH3, CO2, β-аланина и β-аминоизобутирата.
Экзаменационный билет №14
1. Синтез глицерофосфолипидов. Схема синтеза. Липопротеины высокой плотности: формирование, функции и метаболизм.
2.Каспазный каскад. Эффекторная фаза.
Задача.
В крови пациента обнаружена повышенная концентрация непрямого билирубина. На основании каких симптомов, выявленных у пациента, врач назначил этот анализ? Назовите возможные причины повышения концентрациинепрямого билирубина в крови. Какие ещё исследования необходимо сделать больному для установления диагноза?
Дисциплина: биохимия
Синтез фосфолипидов
Синтез глицерофосфолипидов (рис.6.51). Лецитин (фосфатидилхолин) и кефалин (фосфатидилэтаноламин) синтезируются во всех клетках организма за исключением эритроцитов. Наиболее интенсивно эти процессы протекают в печени. Существуют два пути образования этих фосфолипидов в клетках. Один из них сберегающий. Он предполагает повторное использование холина в синтезе фосфатидилхолина (ФХ) и этаноламина – в синтезе фосфатидилэтаноламина (ФЭА). Дело в том, что образование холинаde novo очень “дорого” для организма. Суть сберегающего пути синтеза ФХ включает последовательно задержку, активацию и последующий перенос фосфатхолина из ЦДФ-холина на диацилглицерол.
Сберегающий путь синтеза ФЭА аналогичен пути синтеза ФХ. Только в нем используется предварительно образованный этаноламин. Еще один путь образования в клетках ФЭА – декарбоксилирование фосфатидилсерина.
С другой стороны, ФХ может образовываться из ФЭА. При этом на 1 моль синтезируемого ФХ необходимо 3 моля S-аденозилметионина (S-АМ). S-АМ, являющийся источником активных метильных групп, образуется из метионина и АТФ.
Преобладание того или иного пути образования ФХ зависит от наличия в клетке холина. Если содержание его ограничено, ведущее место занимает синтез ФХ из ФЭА через использование S-АМ. Если холина в клетке достаточно много, на первый план выступает образование ФХ de novo, из ЦДФ-холина и ДАГ.
Рис.6.51. Реакции синтеза фосфолипидов
Синтез фосфатидилсерина происходит путем обмена полярных группировок. Аминокислота серин обменивается с этаноламином в составе ФЭА с образованием фосфатидилсерина.
Рис.6.52.Реакции образования фосфатидилинозитола и кардиолипина
Особенностью образования фосфатидилинозита (рис.6.52), в отличие от других, является то, что активированным субстратом является не спирт, а ДАГ. Активированной формой ДАГ является ЦДФ-диацилглицерол. Энергия для реакции синтеза выделяется в результате расщепления пирофосфатной связи.
П ерестройка фосфолипидов.Эти реакции изменяют жирнокислотный состав фосфолипидов. Принцип их прост. Отщепление одной кислоты и присоединение другой. Фосфолипазы А1и А2отщепляют жирные кислоты от С-1 и С-2 в составе остатка глицерола соответственно. В ходе последующего реацилирования в качестве донора жирных кислот выступает ацил-КоА.
Каспазный каскад
Каспазы представляют собой цистеиновые протеазы, которые расщепляют аминокислотные последовательности после остаткааспарагиновой кислоты. Каспазы образуются за счёт активации прокаспаз (молекулярная масса 32-56 кДа), в составе которых выделяют 3 домена: регуляторный N-концевой домен (продомен), большую (17-21 кДа) и малую (10-13 кДа) субъединицы. Активация происходит путём протеолитического процессинга: все три домена расщепляются, отделяется продомен, а оставшиеся большая и малая субъединицы ассоциируются, образуя гетеродимер. Два гетеродимера в дальнейшем формируют тетрамер — полноценную каспазу с двумя каталитическими участками.
Каспазы обнаружены во большинстве живых организмов. У млекопитающих идентифицировано 13 каспаз.Часть из них в апоптозе не участвует (-1, -4, -5, -11, -13). Остальные каспазы, которые участвуют в апоптозе, разделяют на инициаторные (-2, -8, -9, -10, -12) и эффекторные (-3, -6, -7). Инициаторные каспазы активируют эффекторные каспазы, которые в свою очередь провоцируют и непосредственно участвуют в трансформации клетки. В итоге морфологические и биохимические изменения приводят к гибели клетки по типу апоптоза.
Одна из основных функций эффекторных каспаз заключается в прямом и опосредованном разрушении клеточных структур. Гидролизу подвергаются белки ядерной ламины, разрушается цитоскелет, расщепляются белки, регулирующие клеточную адгезию. Другой важной функцией эффекторных каспаз является инактивация белков, блокирующих апоптоз. В частности расщепляется ингибитор DFF (англ. DNA fragmentation factor — «фактор фрагментации ДНК»), препятствующий активации апоптозной ДНКазы CAD (англ. caspase-activated DNase — «ДНКаза, активируемая каспазами»). Разрушению подвергаются и антиапоптозные белки семейства Bcl-2. Наконец, в результате действия эффекторных каспаз происходит диссоциация регуляторных и эффекторных доменов, участвующих в репарации ДНК, мРНК-сплайсинга и ДНК-репликации.
Экзаменационный билет №15
1.Переваривание белков в желудочно-кишечном тракте. Биологическое значение переваривания. Схема процесса. Характеристика пищеварительных ферментов.
2. Репликация РНК. Характеристика. Этапы. Ферменты.
Задача.
Болезнь Паркинсона – заболевание, характеризующееся повышенным тонусом скелетных мышц (дрожанием конечностей), иногда ограниченностью в движениях. При этом заболевании нарушен обмен дофамина, концентрация которого резко снижается. Для лечения применяют производные ДОФА или ингибиторы МАО. Объясните действие этих препаратов.
Дисциплина: биохимия
Переваривание белков и пептидов в ЖКТ происходит под действием пищеварительных соков, содержащих ферментыпротеазы, которые относятся к классу гидролаз.
Протеазыгидролизуют пептидных связей в белках и пептидах, их делят напротеиназы(эндопептидазы) ипептидазы(экзопептидазы).
Протеиназы(эндопептидазы) катализируют расщепление внутренних пептидных связей в белках и пептидах.
Пептидазы (экзопептидазы)отщепляют от молекул белков и пептидов по одной аминокислоте с карбоксильного или аминного конца. Соответственно различаюткарбоксипептидазыиаминопептидазы. Экзопептидазы функционируют в тонкой кишке.
Дипептидазыгидролизуют дипептиды.
Рекомендуемые страницы:
Воспользуйтесь поиском по сайту:
Источник